Генетика развития подтверждает общее происхождение выделительных органов разных групп животных
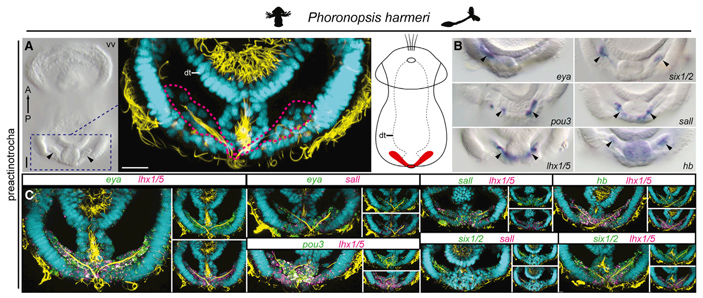
Рис. 1. Экспрессия регуляторных генов в протонефридиях планктонной личинки форониды Phoronopsis harmeri (см. Предок мшанок был «протофоронидой», «Элементы», 03.04.2018). Наверху в центре — прорисовка общего строения личинки, где протонефридии залиты красным; dt — пищеварительный тракт. А — строение области протонефридиев под электронным микроскопом. Вид с брюшной стороны (vv). Голубым цветом окрашены места, где много клеточных ядер, желтым — где много микротрубочек, а пурпурным пунктиром обведены концевые рабочие клетки протонефридиев (циртоциты). B — иммунохимическим методом показана экспрессия в выделительных органах нескольких регуляторных генов, в том числе не упомянутого в тексте гена hb (hunchback), который участвует в развитии выделительных органов только у первичноротых. С — данные об экспрессии детализированы, показана совместная экспрессия разных генов попарно. Форонида выбрана только для примера, в обсуждаемой статье есть такие же данные еще для пяти совершенно разных животных (см. текст). Иллюстрация из обсуждаемой статьи в Current Biology
Выделительные органы многоклеточных животных очень разнообразны. Еще в XIX веке их разделили на протонефридии и метанефридии (не считая некоторых существенных исключений: например, выделительные органы насекомых не относятся ни к тем, ни к другим). Но что было раньше? Имеют ли протонефридии и метанефридии общее происхождении? Если да, то какой тип органов древнее? На эту тему высказывались и высказываются очень разные мнения. Авторы недавних работ по генетике развития дают свой ответ: да, все нефридии имеют единое происхождение, протонефридии появились раньше, метанефридии произошли от них.
Земная жизнь неразрывно связана с водой. Вода — несомненно, самый распространенный в Солнечной системе талассоген (этим словом Айзек Азимов предложил называть вещество, способное сформировать планетарный океан). Может быть, где-то во Вселенной есть жизнь, возникшая на основе других талассогенов — например, аммиака, углекислоты или жидких углеводородов, из которых состоят реки и озера Титана. Но для земной жизни единственная среда, в которой ее интимные механизмы могут функционировать — это водный раствор.
Состав водного раствора отличается внутри живой клетки и за ее пределами. Например, внутри клетки, как правило, гораздо больше ионов калия, чем ионов натрия, а снаружи — наоборот. Эти и подобные различия регулируются сложной системой взаимодействий, которая на профессиональном жаргоне биологов называется «водно-солевой обмен».
Любая живая клетка окружена мембраной, которая обладает свойством полупроницаемости. Это означает, что одни молекулы сквозь нее проходят, а другие нет. Молекулы воды проходят через мембрану очень легко, а крупные органические молекулы не проходят совсем. Между тем любая клетка полна органических молекул, в превращениях которых, собственно, и заключается ее жизнь. Если поместить клетку в дистиллированную воду, в которой вообще нет никаких растворенных веществ, то вода по законам физики устремится из внешней среды (где концентрация воды равна 100%) внутрь клетки (где концентрация воды заведомо меньше 100%, ведь там растворено и что-то другое). Животная клетка, у которой, в отличие от растительной, мембрана не защищена прочной клеточной стенкой, в этом случае просто лопнет.
А что будет, если поместить живую клетку в такой раствор, где растворенных веществ (например, соли или сахара) гораздо больше, чем внутри клетки? Тогда вода устремится из клетки наружу, и клетка погибнет от обезвоживания, несмотря на то, что будет погружена в воду. На этом принципе основано консервирование продуктов — засахаривание или засолка.
Раствор, в котором общая концентрация растворенных веществ ниже, чем в клетке, называется гипотоническим, а раствор, где концентрация растворенных веществ выше — гипертоническим. Растворы, где эти концентрации равны, называются изотоническими. Пример — применяемый в медицине физиологический раствор, в который добавлены соли, чтобы он был изотоничен плазме крови.
В гипотоническом растворе клетка рискует лопнуть, в гипертоническом — высохнуть. Гипотоническим раствором для любой живой клетки является пресная вода. Нас это может удивить, но для земных живых клеток пресная вода — гораздо более чуждая и агрессивная среда, чем морская. Поэтому у пресноводных одноклеточных организмов, особенно у тех, у кого нет клеточной стенки, обязательно появляется сократительная вакуоль, задача которой — постоянно выкачивать из клетки поступающую туда воду. Это прототип выделительной системы.
У животных клеточной стенки не бывает никогда. Поэтому проблемы водно-солевого обмена для них чрезвычайно критичны. Это относится не только к обитателям пресных вод (хотя к ним — в первую очередь). Первое, что должна «научиться» эффективно делать выделительная система, — это выгонять из организма воду. От других ненужных веществ можно избавиться и иначе: например, аммиак у водных животных прекрасно выводится через жабры или даже просто через поверхность тела. Но не вода. Для пресноводных животных умение выводить лишнюю воду — без преувеличения вопрос жизни или смерти, но и морским это бывает полезно, учитывая, что соленость морей может колебаться.
Ионный состав тела животного всегда существенно отличается от состава морской воды, даже если оно изотонично этой воде (что бывает у беспозвоночных довольно часто). Поэтому регулировать концентрацию ионов животному необходимо. Это еще одна задача выделительной системы. И, конечно, выделительная система всегда так или иначе занимается удалением из тела конечных продуктов метаболизма. Это может быть аммиак, мочевина, мочевая кислота или другие вещества, содержащие азот, которые образуются в процессе распада белков.
Почки и не только
У первых многоклеточных животных никаких специальных выделительных органов не было. Губки, гребневики, пластинчатые, стрекающие обходятся без них до сих пор. Губки и стрекающие бывают пресноводными, но в этом случае им хватает сократительных вакуолей в каждой клетке. Настоящая выделительная система — изобретение исключительно двусторонне-симметричных животных, или билатерий.
Типичные выделительные органы билатерий — трубочки, которые называются нефридиями. Жидкость прокачивается через трубочку и в конце концов выводится наружу под действием бьющихся жгутиков или ресничек. Этот процесс называют фильтрацией. Кроме того, выстилающие трубочку клетки дополнительно выделяют в нее вещества, от которых организму надо избавиться (секреция) и всасывают обратно вещества, которые могут еще пригодиться (реабсорбция). Все эти функции есть и в наших почках, хотя механизм фильтрации там другой.
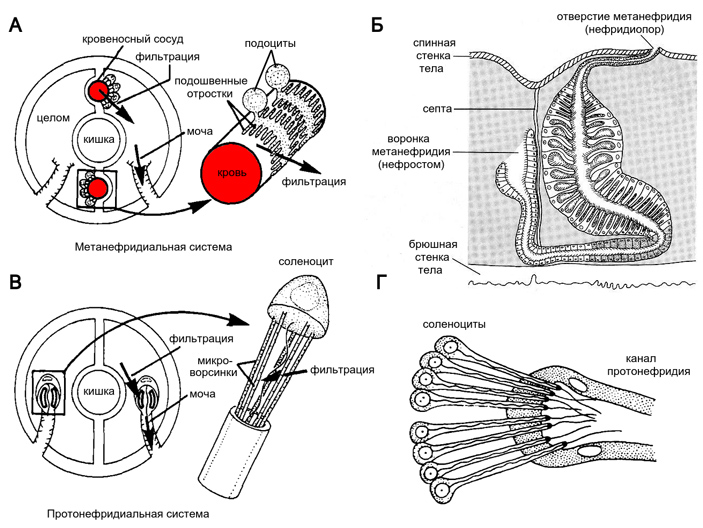
Рис. 2. Метанефридиальная (А, Б) и протонефридиальная (В, Г) выделительные системы беспозвоночных. В метанефридиальной системе жидкость сначала профильтровывается из крови в целомическую полость через щели между отростками клеток целомической выстилки (подоцитов), а потом всасывается в ресничную воронку метанефридия. В протонефридиальной системе полостная или тканевая жидкость профильтровывается через щели между микроворсинками концевой клетки протонефридия (соленоцита), после чего жгутик гонит ее наружу. А, В — иллюстрации из статьи E. Ruppert, P. Smith, 1988. The functional organization of filtration nephridia. Б — метанефридий самца морского кольчатого червя Polydora (S. Rice, 1980. Ultrastructure of the male nephridium and its role in spermatophore formation in spionid polychaetes (Annelida)). Септой называется перегородка между целомами разных сегментов. Мешкообразные впячивания в дальней от воронки части метанефридия — это половые железы, к выделительной функции они прямого отношения не имеют. Г — одно из окончаний протонефридия морского кольчатого червя Phyllodoce (В. А. Догель, 1981. Зоология беспозвоночных)
Австрийский зоолог Бертольд Гатчек (Berthold Hatschek) разделил нефридии на две категории (рис. 2; B. Hatschek, 1888. Lehrbuch der Zoologie). Протонефридий представляет собой вдающуюся глубоко в тело трубочку, замкнутую на внутреннем конце как бы насаженной на нее крупной клеткой, которая высовывает в полость трубочки либо жгутик, либо целый пучок ресничек. Концевая клетка связана с трубочкой микроворсинками, сквозь щели между которыми фильтруется вода. Общее название этих клеток — циртоциты. Одножгутиковые циртоциты называются соленоцитами, а обладающие целым пучком ресничек — пламенными клетками, потому что биение ресничек под микроскопом похоже на пламя (E. Ruppert, P. Smith, 1988. The functional organization of filtration nephridia). Протонефридии часто встречаются у личинок, но иногда и у взрослых животных, например у плоских червей.
Метанефридий состоит из выстланной ресничными клетками воронки, которая открывается в целомическую полость, и трубочки, выводящей жидкость наружу. По самой своей конструкции метанефридий мощнее протонефридия: ведь на внутреннем конце трубочки протонефридия циртоциты сидят поодиночке, а в метанефридии ресничных клеток сразу много, и они работают сообща. Трубочка метанефридия часто бывает извитой, по ее ходу активно идут секреция и реабсорбция, так что вторичная моча, которую животное выводит во внешнюю среду, может сильно отличаться по составу от целомической жидкости. Метанефридии характерны, например, для кольчатых червей. У животных, не имеющих целома, их по определению быть не может.
У ланцетника нефридии вдаются внутренним концом в целом, но при этом оканчиваются типичными циртоцитами (рис. 3). Протонефридии это или метанефридии? Решение этого вопроса зависит от того, какой критерий считать определяющим. Дело в том, что целом у ланцетника очень маленький. Он вытеснен огромной околожаберной (атриальной) полостью, в которую нефридии, кстати говоря, и впадают. Объем оставшегося целома так мал, что циртоциты, вдающиеся в целомическую камеру, упираются в ее противоположную стенку, да еще и заякориваются на ней с помощью специальных подошвенных отростков.
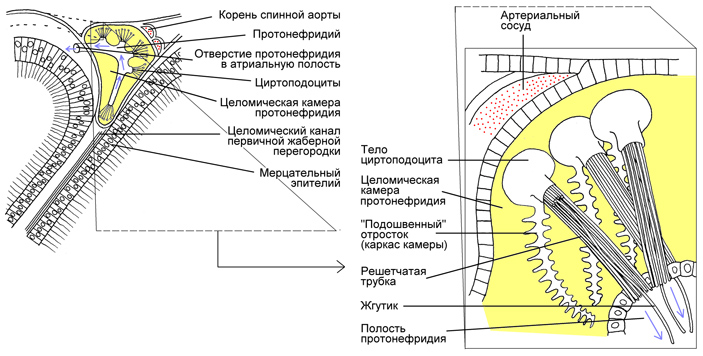
Рис. 3. Выделительная система ланцетника. Желтым цветом залит наджаберный целом. Голубые стрелки обозначают направление тока жидкости в нефридии. Решетчатая трубка образована микроворсинками. Точное название жгутиковых клеток нефридиев ланцетника — циртоподоциты: они сочетают особенности классических циртоцитов и подоцитов целомической выстилки (см. далее). Другие пояснения в тексте. Иллюстрация из книги Ф. Я. Дзержинский, 2005. Сравнительная анатомия позвоночных животных
С легкой руки авторитетного английского биолога Эдвина Гудрича (Edwin Stephen Goodrich) выделительные органы ланцетника стали считать протонефридиями (см. N. Holland, 2017. The long and winding path to understanding kidney structure in amphioxus — a review). Но классические протонефридии имеют эктодермальное происхождение: они развиваются как глубокие впячивания наружного покрова. А у ланцетника, как показала электронная микроскопия, циртоциты представляют собой сильно модифицированные клетки целомической выстилки, в состав которой они, собственно, и входят (T. Stach, K. Eisler, 1998. The ontogeny of the nephridial system of the larval amphioxus). Так что по происхождению они родственны ресничным клеткам метанефридиев. Сходство с протонефридиями возникло из-за крайнего уменьшения целома, которое позволило разместить жгутиковые клетки не внутри входа в нефридий, а напротив него.
У позвоночных атриальной полости нет. Целом у них большой, и выделительная система напоминает обычные для целомических животных метанефридии. Но — с важным дополнением. Если у беспозвоночных выделительные трубочки обычно открываются наружу по отдельности (например, посегментно), то у позвоночных они сливаются в общий проток, образуя почку. Нефридии, включенные в состав почки, называются нефронами.
Кроме того, у позвоночных появилось сердце — центральный мышечный насос, качающий кровь по сосудам под высоким давлением. Этого давления достаточно, чтобы нагнетать жидкость из кровеносных сосудов прямо в нефроны. Такой источник энергии было бы невыгодно оставлять в стороне, поэтому почки позвоночных всегда тесно связаны с кровеносной системой. У позвоночных, имеющих так называемую головную почку, рядом с каждой воронкой нефрона находится почечный клубочек — компактное переплетение тонких кровеносных сосудов, откуда под давлением просачивается жидкая составляющая крови. А у позвоночных, имеющих туловищные и тазовые почки, каждый нефрон образует боуменову капсулу — бокаловидный полый вырост, охватывающий сосудистый клубочек почти со всех сторон (рис. 4). Клетки целомического эпителия боуменовой капсулы называются подоцитами. Они имеют примерно такие же подошвенные отростки, как целомические циртоциты ланцетника, и иногда считаются их гомологами (E. Ruppert, 1994. Evolutionary origin of the vertebrate nephron).
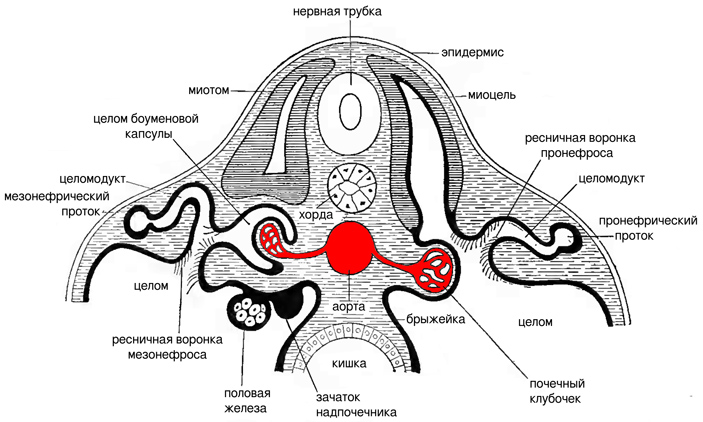
Рис. 4. Нефроны головной (справа) и туловищной (слева) почки на поперечном разрезе зародыша позвоночного. Целом тут очень велик, в частности продолжение целома (миоцель) есть в зачатке осевой сегментарной мускулатуры (миотоме). Стенки правого и левого целомов, сблизившись посредине, образуют брыжейку, в которой проходит аорта. Головная почка здесь названа пронефросом, туловищная — мезонефросом, а извитой каналец нефрона — целомодуктом, потому что он развивается как продолжение целома. Нефроны каждой почки впадают в общий проток, соответственно пронефрический или мезонефрический. И в головной, и в туловищной почке сохраняются ресничные воронки, унаследованные от метанефридиев. Но в туловищной почке, кроме того, есть боуменова капсула — бокаловидный вырост целома, охватывающий сосудистый почечный клубочек, чтобы сделать фильтрацию более эффективной. Иллюстрация из книги R. Wiedersheim, 1907. Comparative anatomy of vertebrates
По пути из сосуда в нефрон кровь фильтруется дважды: через эпителий сосуда и через эпителий боуменовой капсулы, где между звездчатыми подоцитами есть узкие щели. Ни клетки крови, ни плавающие в ней молекулы белков сквозь эти фильтры в норме не проходят. Остается плазма крови, лишенная белков, которая скапливается в полости боуменовой капсулы и называется первичной мочой. Во вторичную мочу она превращается после того, как пройдет через извитой каналец нефрона и подвергнется процессам реабсорбции и секреции.
В боуменовой капсуле фильтрация идет без помощи ресничных клеток, исключительно за счет давления крови. В туловищных почках рядом с боуменовыми капсулами могут сохраняться унаследованные от метанефридиев ресничные воронки, но в тазовых почках их нет. У человека, как и у всех млекопитающих, почка тазовая. Вся фильтрация в ней идет через боуменовы капсулы.
Нужно пояснить, что головные, туловищные и тазовые почки — это эмбриональные поколения почек, которые могут сменять (а могут и не сменять) друг друга в развитии позвоночных. Принято считать, что головная почка сохраняется во взрослом состоянии только у миксин. С этим связан один забавный литературный казус. В романе японского писателя Кобо Абэ «Четвертый ледниковый период», который перевел на русский язык Аркадий Стругацкий, можно прочитать: «Передние почки у взрослого организма имеются только у угрей». Это, несомненно, ошибка, но чья? В оригинале романа использовано японское слово, обозначающее миногу (но не миксину). В английском переводе фигурирует тоже минога. Таким образом, тут наложились друг на друга две неточности: сначала Кобо Абэ перепутал миксину с миногой, а потом Аркадий Стругацкий перепутал миногу с угрем. Редкий для Стругацких пример переводческой ошибки.
У миног, рыб и амфибий почка туловищная, а у рептилий, птиц и млекопитающих — тазовая. Этот факт тоже нашел отражение в художественной литературе. В повести Михаила Булгакова «Роковые яйца» профессор Персиков говорит студенту: «Как, вы не знаете, чем отличаются голые гады от пресмыкающихся? Это просто смешно, молодой человек. Тазовых почек нет у голых гадов. Они отсутствуют. Так-то-с». Такой разговор действительно мог бы произойти в начале XX века в одной из аудиторий Московского университета — Булгаков знал, о чем пишет.
Нефридии, черви и гены
Итак, нефридии (или их производные) присущи большинству крупных групп билатерий, от плоских червей до позвоночных включительно. Наличие нефридиев служит общим признаком двух величайших эволюционных ветвей животного царства — первичноротых и вторичноротых. По этой причине шведский зоолог Ульф Йонделиус и его коллеги предложили объединить первичноротых и вторичноротых в группу Nephrozoa (U. Jondelius et al., 2002. The Nemertodermatida are basal bilaterians and not members of the Platyhelminthes). По-русски ее можно было бы назвать «нефрозои», животные с нефридиями.
Однако не все билатерии входят в число нефрозоев. Есть билатерии, не относящиеся ни к первичноротым, ни к вторичноротым: это ксенотурбеллиды, бескишечные турбеллярии и немертодерматиды — своеобразные морские черви, которых объединяют в группу Xenacoelomorpha. Группа эта немногочисленна, но проигнорировать ее существование никак нельзя. Предполагается, что ксенацеломорфы — древнейшая эволюционная ветвь билатерий, обособившаяся раньше, чем первичноротые и вторичноротые разделились (см. Ксенотурбеллиды оказались близки к предкам двусторонне-симметричных животных, «Элементы», 15.02.2016). Никаких нефридиев у них нет.
В последнее время, правда, дискуссия вокруг ксенацеломорф в очередной раз оживилась. Появились молекулярные данные, согласно которым ксенацеломорфы на самом деле близки к вторичноротым (см. А существуют ли вторичноротые?, «Элементы», 31.05.2021). Эту гипотезу высказывали и раньше, но сейчас она получила новую поддержку. Если она верна, то Bilateria и Nephrozoa — просто синонимы.
Так или иначе, этот вопрос пока не решен. Более того, неизвестно, возможно ли тут вообще окончательное решение: в случае с ксенацеломорфами молекулярная филогенетика уже почти исчерпала свои ресурсы, которые очень велики, но не беспредельны. Это добавляет неопределенности в запутанную историю выделительных органов животных, где, в свою очередь, хватает нерешенных проблем. Что появилось раньше — протонефридии или метанефридии? Какие выделительные органы были у первых билатерий? Появлялись ли нефридии много раз независимо, или все они имеют общее происхождение?
В подобных случаях у современных исследователей часто возникает соблазн бросить на чашу весов некий решающий пакет молекулярных данных, истолкование которого расставит все по местам раз и навсегда. А эволюционная биология развития дает немало возможностей, наводящих на такие мысли.
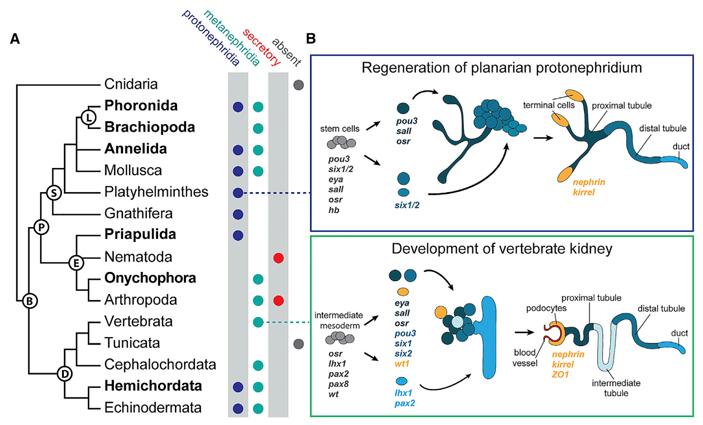
Рис. 5. Выделительные органы в разных ветвях билатерий (A) и их молекулярная общность (B). Обозначения эволюционных ветвей: B — Bilateria, D — Deuterostomia, P — Protostomia, E — Ecdysozoa, S — Spiralia, L — Lophotrochozoa. Ксенацеломорфы не показаны. Почки позвоночных здесь отнесены к метанефридиям, и выделительные органы ланцетника — тоже к метанефридиям, в отличие от книги Ф. Я. Дзержинского, из которой взят рис. 3. У некоторых групп, например у кольчатых червей и у моллюсков, встречаются как метанефридии, так и протонефридии. Кроме того, есть особая (не обсуждаемая в тексте) категория выделительных органов, которые вообще не относятся к нефридиям, потому что фильтрации в них нет — только секреция. Такие чисто секреторные органы выделения характерны для нематод и для членистоногих. В обсуждаемом исследовании они не рассматривались. B — экспрессия одних и тех же генов (и регуляторных, и структурных) в развивающихся протонефридиях плоских червей и нефронах позвоночных. Протонефридий и нефрон нарисованы так, чтобы показать сходство. Иллюстрация из обсуждаемой статьи в Current Biology
Итак, лет десять назад было замечено, что некоторые гены, кодирующие факторы транскрипции, экспрессируются, с одной стороны, в ходе развития почек позвоночных, а с другой — при регенерации протонефридиев плоских червей (рис. 5). Ясно, что более далеких друг от друга животных не найти, по крайней мере если оставаться в пределах нефрозоев. Как объяснить активность одних и тех же генов в пламенных клетках планарии и в подоцитах боуменовой капсулы мыши?
Основываясь на этих фактах, группа ученых провела исследование экспрессии генов в развитии выделительных структур нескольких животных — причем как можно более разных, чтобы получить полную картину. В число объектов вошли такие относительно малоизвестные существа, как форониды (рис. 1), брахиоподы, приапулиды, онихофоры и полухордовые.
В результате удалось выделить группу как минимум из пяти регуляторных генов (eya, six1/2, pou3, lhx1/5, osr), которые экспрессируются в клетках развивающихся выделительных органов и у первичноротых, и у вторичноротых. Четыре гена (eya, six1/2, pou3, lhx1/5) обладают этой функцией у всех исследованных животных без исключения. Связанная с выделительными органами экспрессия гена osr, видимо, исчезла у кольчатых червей, форонид и брахиопод, но сохранилась у плоских червей, которые тоже относятся к ветви спиральнодробящихся. Кроме того, она отсутствует у полухордовых, зато присутствует у позвоночных. В общем, есть гены, которые ведут себя в эволюции как устойчивый «нефридиальный комплекс».
Эти данные касаются регуляторных генов, продукты которых могут служить более или менее универсальными переключателями. Все они могут брать на себя и другие функции, кроме участия в развитии выделительной системы. Но есть еще и структурные гены, кодирующие белки, которые физически обеспечивают работу выделительных органов. Например, трансмембранный белок нефрин (см. Nephrin) у человека сидит на подоцитах боуменовой капсулы, помогая формировать щели, через которые идет фильтрация. Но тот же ген нефрина экспрессируется и в пламенных клетках протонефридиев плоского червя! А ведь почка человека и протонефридий планарии на первый взгляд ничем друг на друга не похожи.
Широкое исследование показало, что экспрессия гена nephrin и еще двух генов, близких к нему по функции, наблюдается в нефридиях или почках почти у всех животных, у кого эти органы вообще есть. А найденные исключения, пожалуй, только подтверждают правило. Например, у птиц, в отличие от других позвоночных, нефрин в работе почек не участвует — факт, который немало удивил физиологов (J. Milner, 2012. Life without nephrin: it’s for the birds). Но это наверняка эволюционно недавняя утрата.
О чем все эти открытия свидетельствуют?
Авторы недавно вышедшей большой статьи — а среди них есть очень известные биологи, такие как Андреас Хейноль (Andreas Hejnol), Грэм Бадд (Graham Budd) и Кристофер Лоу (Christopher Lowe) — за этими тремя людьми числятся великолепные работы по зоологии, палеонтологии и биологии развития, — так вот, авторы статьи делают естественный вывод, что обнаруженные ими факты свидетельствуют о гомологии всех типов нефридиев. Это должно означать, что у общего предка нефрозоев (если не всех билатерий) нефридии уже были.
Ну, и что же это были за нефридии? Ответ: протонефридии. Во-первых, из полученных данных видно, что экспрессия генов «нефридиального комплекса» наиболее полно — можно сказать, классически — проявляется именно в протонефридиях: у планктонных личинок морских кольчатых червей и форонид, у взрослых плоских червей и приапулид. Во-вторых, есть чисто морфологические работы, без всяких генов изящно обосновывающие гипотезу, что протонефридии — исходные для билатерий органы выделения. У маленьких животных формируются протонефридии, а у крупных метанефридии (E. Ruppert, P. Smith, 1988. The functional organization of filtration nephridia). (Последнее, правда, при условии, что у них есть целом. Даже самый крупный плоский червь будет вынужден обойтись протонефридиями, ибо у него целома нет.)
Наконец, хотя в статье это и не упомянуто, предположение о первичности протонефридиев хорошо соответствует биогенетическому закону, согласно которому развитие особи, как правило, кратко повторяет эволюцию. Есть много животных, у которых личинки имеют протонефридии, а взрослые формы — метанефридии. Наоборот не бывает.
Авторы не исключают, что метанефридии могли произойти от протонефридиев несколько раз, независимо в разных группах животных. Такая идея существует давно (T. Bartolomaeus, P. Ax, 1992. Protonephridia and metanephridia — their relation within the Bilateria), и новые данные ей не противоречат. Тогда метанефридии разных животных совсем не обязательно должны быть гомологичны друг другу. Точнее, они могут быть гомологичны друг другу в качестве органов выделения, но негомологичны именно в качестве метанефридиев, подобно тому, как крыло птицы и крыло летучей мыши гомологичны друг другу в качестве тетраподной конечности, но негомологичны в качестве крыльев (пример не выдуманный, а предложенный в той же статье).
Еще на один аспект проблемы нефридиев обратил внимание видный знаток эволюционной биологии развития животных Детлев Арендт (Detlev Arendt). Дело в том, что из пяти регуляторных генов «нефридиального комплекса» как минимум два (eya, six1/2) участвуют еще и в развитии внутреннего уха позвоночных. Почему это важно? А потому что чувствительные клетки внутреннего уха построены по той же модели, что и соленоциты примитивных протонефридиев: на вершине клетки сидит жгутик, а рядом с ним — группа микроворсинок. И эта модель, судя по данным генетики развития, унаследована от воротничковых жгутиконосцев — ближайших одноклеточных родственников животных, клетки которых устроены очень похоже (см. «Первичность губок» опережает по очкам «первичность гребневиков», «Элементы», 12.11.2019). Более того, в соленоцитах «воротничковая» модель клетки сохранена полнее, чем в клетках внутреннего уха: микроворсинки там не располагаются группой в стороне от жгутика, а окружают его кольцом — как у жгутиконосцев. Теперь есть новые поводы считать, что это сходство не случайно. Оно отражает очень глубокую эволюционную преемственность.
Нерешенные вопросы
Полученные результаты отлично иллюстрируют общую закономерность эволюции эукариот: сложные морфологические структуры складываются из устойчивых молекулярных «кирпичиков», возникших давным-давно, когда их обладатели были устроены куда более просто. Множество иллюстраций можно найти, например, в истории эволюционного становления нервной системы: чем больше там открывается новых фактов, тем этот принцип яснее (см. У губок найдены вероятные эволюционные предшественники нейронов и миоцитов, «Элементы», 08.11.2021). Кстати говоря, белок нефрин используется не только в выделительной, но и в нервной системе — для установления контактов между нейронами. Участвует он и в формировании кровеносных сосудов (N. Wagner et al., 2011. The podocyte protein nephrin is required for cardiac vessel formation). Это тоже закономерно: один и тот же молекулярный «инструмент» допускает различные применения.
Все это прекрасно, но остаются и проблемы, до решения которых сюжет эволюции выделительной системы никак нельзя считать исчерпанным.
Во-первых, теория первичности протонефридиев игнорирует эмбриологию. Сравнительные анатомы беспозвоночных всегда утверждали, что протонефридии образуются из эктодермы (В. Н. Беклемишев, 1964. Основы сравнительной анатомии беспозвоночных. Т. 2). Это означает, что они имеют совсем иное происхождение, чем метанефридии, основой которых служат целомические воронки. Объяснить это легче всего, предположив, что протонефридии и метанефридии возникли в ходе эволюции независимо друг от друга: метанефридии — у взрослых целомических животных, а протонефридии — у планктонных личинок (см. такую интерпретацию в статье: В. В. Малахов, Е. В. Богомолова, 2016. Новый взгляд на строение и происхождение полости тела многоклеточных). Более того, эта идея отлично вписывается в сценарий происхождения планктонных личинок билатерий, который недавно предложили академик Владимир Васильевич Малахов и его коллеги (см. Общее происхождение трохофор и диплеврул: за и против, «Элементы», 06.02.2020). Ну а как тогда объяснить наличие протонефридиев у взрослых организмов? Очень просто: неотенией, размножением на личиночной стадии. В этом случае протонефридии сохраняются, а у крупных животных и усложняются, подобно перистым жабрам у аксолотля. Сторонники единства нефридиев либо вообще отмахиваются от всех этих соображений (E. Ruppert, P. Smith, 1988. The functional organization of filtration nephridia), либо ссылаются на данные по кольчатым червям и полухордовым, у которых вроде бы описаны переходные состояния между протонефридиями и метанефридиями (см. обсуждаемую статью Арендта). Но прояснить этот вопрос определенно надо.
Во-вторых, остается нерешенной проблема ксенацеломорф. У них тоже обнаружена экспрессия генов «нефридиального комплекса», в том числе и гена nephrin, от которого зависит работа нефронов позвоночных (C. Andrikou et al., 2019. Active mode of excretion across digestive tissues predates the origin of excretory organs). При этом нефридии у ксенацеломорф полностью отсутствуют, а аналогичные функции у них, похоже, выполняют пищеварительные клетки. Таким образом, экспрессия генов «нефридиального комплекса» не равносильна наличию нефридиев как морфологических структур. Ничего неожиданного тут нет — эволюционная биология развития то и дело сталкивается с такой проблемой (см. У одноклеточных организмов есть ген, способный управлять развитием хорды, «Элементы», 25.10.2013). Судить по генам о морфологических структурах можно лишь осторожно и опосредованно, прямолинейность в этом вопросе часто ведет к ошибкам (см. C. Nielsen, P. Martinez, 2003. Patterns of gene expression: homology or homocracy?). Хейноль, Бадд и Лоу с коллегами теперь считают, что предки ксенацеломорф когда-то имели настоящие нефридии, но потом потеряли их. Очевидно, это либо верно, либо нет. Здесь тоже есть что исследовать.
Источники:
1) Ludwik Gasiorowski, Carmen Andrikou, Ralf Janssen, Paul Bump, Graham E. Budd, Christopher J. Lowe, Andreas Hejnol. Molecular evidence for a single origin of ultrafiltration-based excretory organs // Current Biology. 2021. DOI: 10.1016/j.cub.2021.05.057.
2) Detlev Arendt. Animal evolution: of flame and collar cells // Current Biology. 2021. DOI: 10.1016/j.cub.2021.07.006.
Сергей Ястребов